 Little Gull Larus minutus
Little Gull Larus minutus
(last update: March 02, 2015)
Little Gull adult
IDENTIFICATION OF LITTLE GULL Larus minutus
(from: Malling Olsen, K. & Larsson, H. 2004 Gulls of Europe, Asia and North America.
London: Christopher Helm).
PART 4
GEOGRAPHICAL VARIATION
Not fully documented. Breeding males from Siberia on average have longer wings than in other populations (mean 230.6), but sample restricted.
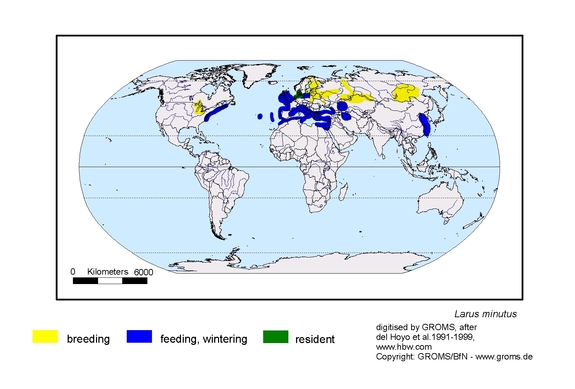
DISTRIBUTION AND MIGRATION
Probably some tens of thousands of pairs, mainly in former USSR. Breeds on grassy islands, sandbanks and reedbeds in fresh shallow lowland lakes, often together with Black-headed Gull and marsh terns. In E Baltic in brackish water. On migration along shores, on reservoirs and lagoons, lakes and at sea. Winters at sea and along sandy or muddy beaches (Ill”icev & Zubakin 1990, Hagemeijer & Blair 1997).
European population mid-late 1990s (in pairs) 23,000-32,000: Russia 11,000-14,000; Sweden 500 (90-125 early 1990s); Norway max. 10 (first bred 1976); Finland 8,000-10,000 (first bred 1879; strong increase in recent decades); Poland 20-100 (most Lake Druzno); Latvia 700-3,000; Estonia 1,000-2,000; Belarus 1,800-2,300; Lithuania 100-200; Ukraine 100-200. Netherlands 5-40 (61) (first bred 1942, after which irregular and often unsuccessful, but regular 1975-1988 on Lauwersmeer; since then most records Waddensee and Delta area). Irregular Great Britain, Germany, Denmark (1940-1950s >50-75 pairs W/N Jutland; since 1980 occasional single pairs), France, C Europe and Romania. General decrease in S part of breeding range but increase in N Sweden and Baltic since 1970 (Cramp 8c Simmons 1983, Tucker & Heath 1994, del Hoyo et al. 1996, Hagemener & Blair 1997, Svensson et al. 1999, Bijlsma et al. 2001).
Recently spread to N America. First record 1919-1922, since 1987 regular. First bred Canada 1962, USA 1975; subsequent breeding records in many states (Glutz von Blotzheim & Bauer 1982, Godfrey 1986, del Hoyo et al. 1996).
Migratory. Arrives breeding sites late Apr-late May, leaves from late Jul. Population from Fennoscandia and W Russia migrates W and SW to main wintering grounds W Mediterranean and Atlantic coast N to Brittany; Finnish and Baltic birds recovered W to Great Britain and E to Adriatic / Black Sea, extralimitally Kazakhstan.
In autumn usually coastal, but thousands (mainly second-years and adults) gather N German lakes and low wetlands to moult before further migration; leaves early-mid-Sep. Migration in N and W Europe: large migration S Baltic, to a lesser degree Swedish Great Lakes (mainly first-years). Large migration Swedish and Danish Baltic coastline (N to Gotland), where on average 13,000 pass yearly. Most observed from coast in strong E-S winds combined with rain and low visibility. First-years start Jul and peak mid-Aug-mid-Sep. Migration route of northern (Finnish?) populations passes mid-Baltic, max. 1,350 at Bjorn, Uppland, on 16-18 August 1996. Head SW across Swedish lakes (Mälaren-Vänern) to N Kattegat. Adults much scarcer along this route, but flocks of max. 200 noted mid-late Oct. First-years otherwise generally head S across C Europe.
Adult migration in S Baltic starts late Aug, peaks mid-Oct-early Nov. Visible migration greatly dependent on weather; best years around 6,000 Scania and Öland, Sweden, respectively. Best autumns 1994 (with unsurpassed peak mid-Sep) and 2000 following hard SSE winds and poor visibility. In Sweden max. day-counts 3,565 at Öland on 16 August 1994 (31% adult, 8% second-years, 61% first-years), 3,309 at Segerstad on 7 November 2000 and 1,614 at Sandhammaren on 29 October 2000 (1,223 adults, 115 second-years and 148 first-years). At Gedser, SE Denmark, 4,000 in best season (Glutz von Blotzheim & Bauer 1982, Koop 1985, Breife 1987, Jensen 1993, Darefeldt & Johansson 1994, Dahlgren et al. 1995, Lindberg & Lotberg 1997, Arinder 2001, pers. obs.).
Max. 500 Scotland late Aug-Oct suggests arrival via Swedish Great Lakes and N Kattegat. Gathers at moulting sites in N Great Britain before heading W and S late Sep-Oct (see below); situation similar to German. Along Danish W coast regular; day-counts >600 in strong SSW winds. Large passage Dutch W coast and Belgium (late Jul) Aug-mid-Oct (max. 10,000 Netherlands; mainly first-years into early Oct, followed by adults). Peak Oct-Nov; max. day-counts 2,238 Netherlands and 1,435 Zeebrugge, Belgium. Probably arrives after large-sca1e inland migration W from S part of breeding range. Peak of mainly adults Pas-de-Calais, NW France, early Nov. Regular C European lakes. In Bodensee peak late Aug-late Sep. Max. 280, mainly first-years (Hutchinson & Neath 1978, Cramp & Simmons 1983, Schuster et al. 1983, Malling Olsen 1992, Murray 1994).
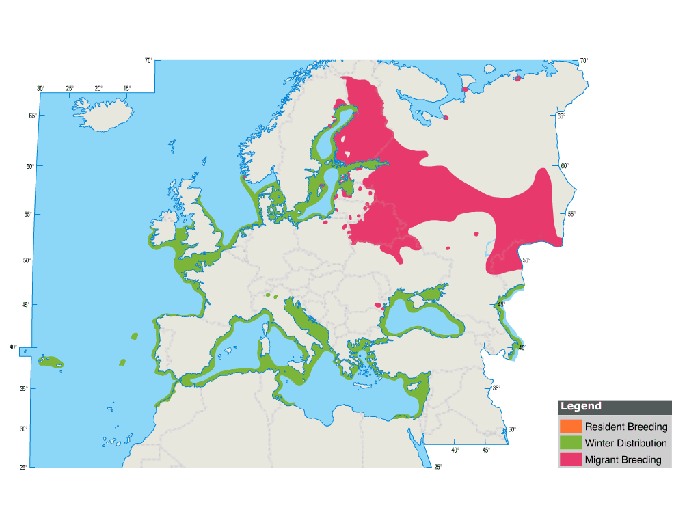
European migrant population heads W for Atlantic Ocean between Irish Sea and Morocco (some further S to Nigeria and Angola) to offshore wintering sites; >600 seen from land in onshore winds Wicklow, Ireland, Jan. British winter population 1,000; largest European winter count 5,000-10,000 Dorset late Dec 1974. Around 1,500 winter Dutch Waddensee (exceptionally 10,000). Scarce Baltic Sea in winter; in Sweden max day-counts 190 Öland and 482 E Scania Dec-Feb, more irregular Swedish Great Lakes in mild winters or before freeze-up; max. 215 Vanersborg on 4 March 1992 (>90% adults) (Glutz von Blotzheim & Bauer 1982, Akerman 1986, Breife 1987, Darefeldt & Johansson 1994, Bijlsma et al. 2001).
In spring, large gathering of adults, Camargue, France (max. 1,560 Apr; in May mainly first-years). At Gibraltar and Cap Gris Nez, France, peak late Mar-late Apr. Of 2,225 Cap Gris Nez 88% adult, 1% second-years, 11% first-years, followed by larger numbers of first-years in May. Large concentrated passage N Netherlands (max. 80,000 in one year) mid-Apr-mid-May; most 4,008 30 April-2 May 1976. Several thousands Apr-May Krammer-Volkerak, SW Netherlands. Along Merseyside coast, NW England, peak second half of Apr (max. day-count 680); >90% leave before mid-May, probably mainly heading for Finland, where migration peak concentrated first weeks May. In spring 1987 large passage (4,136) Heligoland, Germany: peak late Apr-early May; 1,410 on 5 May. Mainly adults in Apr, from late month increasing numbers of first-years: on 5 May 78-80% first-years, 16-18% second-years, 4% adults. Largest German influx noted 30 April 1998: 21,000 Schleswig-Holstein. In Switzerland, max. 1,300 Préverenges on 30 April 1999. These influxes, caused by strong SE winds, also noted Great Britain. Often settles on lakes. Bodensee peak similar timing to above-mentioned, with adults passing late Apr-early May; from late Apr dominance by first- and second-years; of 145 on 5 May only four adults. In S Scandinavia, following SSE winds, flocks of max. 200 late Apr-early May Denmark and Scania (adults; first- and second-years dominant from mid-May). First-years summer in small groups C and N European lakes. Small numbers of all ages summer Great Britain (Woutersen 1980, Glutz von Blotzheim & Bauer 1982, Cramp & Simmons 1983, Schuster et al. 1983, Whittington 1987, Moritz 1988, Messenger 1993, Meininger 1995, Birding World 12: 189, Brit. Birds 94:138, H.J. Lehto in litt., pers. obs.).
C Siberian population migrates via Aral, Caspian and Black Seas to E Mediterranean. Common migrant N and W coast of Black Sea and Turkey (Anatolia, Bosporus, Dardanelles) Aug-Sep. Max. 30,000 Romania, early Aug. Max. counts Bulgaria and Bosporus 2,500 mid-Aug-early Sep. Fewer cross inland Turkey. Some hundreds winter Black Sea and Balik Golu, Turkey; 2,000-5,000 Greece. In Mediterranean intermix with W population. Recovery of bird from Barabinsk, C Siberia, in Belgium (Erard 1960, Cramp & Simmons 1983, Handrinos & Akriotis 1997).
Many winter E Mediterranean. In Nile Delta survey Dec-Jan 1989/ 1990 of 52,769 counted 47,316 were at Lake Manzala 7-8 January 1990 (largest number ever counted). Only a few hundreds there late 1970s and 1980s suggests recent change in winter quarters. Along Israeli Mediterranean coast max. flocks of 460 Jan-Feb, mostly following strong onshore winds. Israeli winter population 2,000-4,000, mainly first-years. In W Mediterranean, max. 1,300 noted off E coast of Spain, 900 Tunisia, hundreds Oran, Algeria and max. 100 Tripoli. Strong movements at Gibraltar during easterly gales. Abundant winterer in Caspian Sea (Bourne 1957, Bundy 1976, Cramp & Simmons 1983, Finlayson 1992, Meininger & Sorensen 1993, Shirihai 1996b).
Spring migration of C Siberian population timed similar to westerly; off Port Said 5,000 late Mar 1990; in E Mediterranean generally largest numbers Apr (max. 2,000 certain Greek sites), sometimes May (e.g. 5,000 Evros Delta, May 1968). Flocks of first-years often summer (Cramp & Simmons 1983, Meininger & Sorensen 1993, Handrinos & Akriotis 1997).
Movements and winter quarters of E population less known; probably winters Seas of Okhotsk and Japan S to China. Erard (1960) suggests that main wintering zone is Sea of Okhotsk. According to Il’icev & Zubakin (1988) China holds three widely separated wintering areas, in NW China, NE China and along the E China Sea coast.
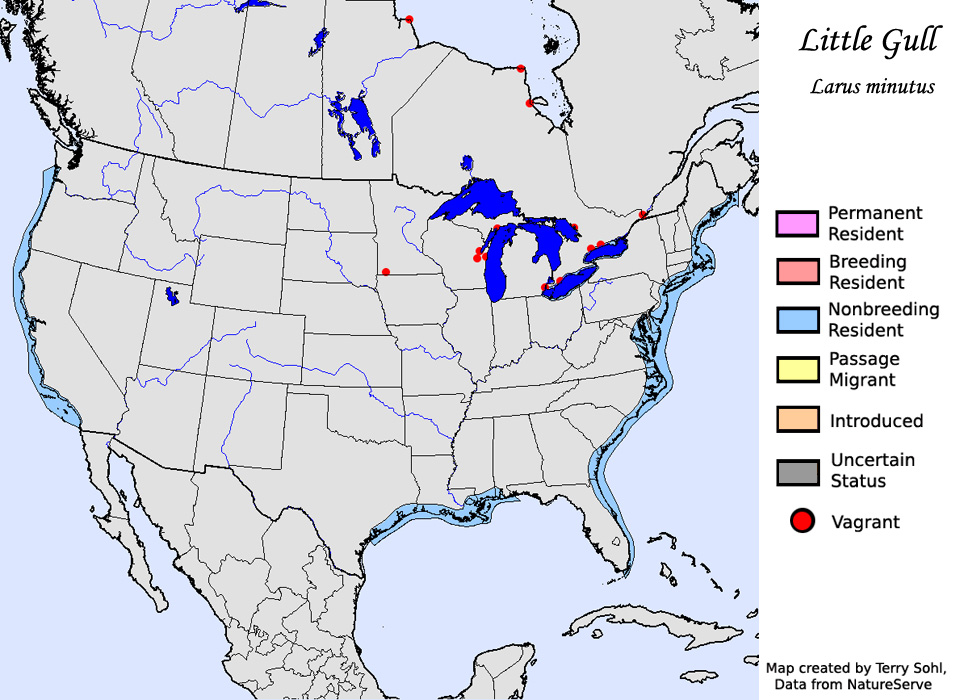
Migration of N American population associated with Bonaparte’s Gull. Max. 25 Niagara Falls/ New England, E Coast, and max. day-count 112, N Carolina, winter and spring, and 50 Lake Ontario, spring. In autumn, max. 10 per day New York, Sep. In winter small numbers between Massachusetts and New York. Vagrant to most states W to California. Autumn arrival to N America probably associated with W movements from W Palearctic. In Iceland 85 into 1992, most May-Jun (first-summer) and Sep (adults). Vagrant Greenland, Arabian Gulf (e.g. seven, UAE), Afghanistan, India, Canary Islands, Azores, Bermuda, Caribbean, tropical Africa S to Sierra Leone, Nigeria and Kenya (largest number 60 Jan 1979 Lake Turkana could indicate unknown winter area) (Cramp & Simmons 1983, Urban et al. 1986, Péturson 1987, Boertmann 1994, del Hoyo et al. 1996).
MEASUREMENTS
Lengths in mm; weights in g. Own data based on skins in NHM, NNH, UZM, ZMA, ZMO. Netherlands, W Europe, Russia, Siberia and Middle East.
WING |
|||
| Adult male | 214-239 | (225.6) | 76 |
| Adult female | 211-236 | (221.5) | 59 |
| First-year male | 215-234 | (220.8) | 44 |
| First-year female | 209-228 | (217.3) | 46 |
TAIL PROFILE (differences between T1 and T6) |
|||
| Adult | 0-5 | (2.2) | 120 |
| First-year | 3-10 | (6.8) | 90 |
BILL |
|||
| Adult male | 21.5-26.7 | (23.2) | 70 |
| Adult female | 21.1-24.2 | (22.9) | 51 |
| First-year male | 20.0-24.0 | (23.0) | 43 |
| First-year female | 19.8-25.1 | (22.0) | 43 |
BILL DEPTH AT BASE OF BILL |
|||
| Adult male | 5.7-7.5 | (6.5) | 69 |
| Adult female | 5.4-7.1 | (6.2) | 54 |
| First-year male | 5.6-7.0 | (6.3) | 44 |
| First-year female | 5.3-6.8 | (6.1) | 42 |
BILL DEPTH AT GONYS |
|||
| Adult male | 4.8-6.0 | (5.3) | 69 |
| Adult female | 4.7-5.9 | (5.2) | 54 |
| First-year male | 4.8-5.8 | (5.2) | 42 |
| First-year female | 4.6-5.4 | (5.0) | 40 |
GONYS LENGTH |
|||
| Adult male | 8.2-10.4 | (9.5) | 69 |
| Adult female | 8.2-10.2 | (9.3) | 52 |
| First-year male | 6.6-10.8 | (9.2) | 45 |
| First-year female | 6.5-10.1 | (8.7) | 41 |
TARSUS |
|||
| Adult male | 23.7-29.0 | (25.8) | 70 |
| Adult female | 23.6-29.4 | (25.2) | 52 |
| First-year male | 22.5-28.8 | (25.7) | 45 |
| First-year female | 23.1-27.0 | (25.1) | 40 |
Weight Adult male 82-127, female 68-133, first-year 66-121 (Glutz von Blotzheim & Bauer 1982, Cramp & Simmons 1983, skins in NNH, UZM, ZMA).
February
 Little Gull minutus, adult, February 09 2014, Wherrytown, UK. Picture: Steve Rogers.
Little Gull minutus, adult, February 09 2014, Wherrytown, UK. Picture: Steve Rogers. April
 Little Gull minutus, adult, April 08 2015, Riga, Latvia. Picture: Igor Deņisov.
Little Gull minutus, adult, April 08 2015, Riga, Latvia. Picture: Igor Deņisov.March
 Little Gull minutus, adult, March 01 2014, Lancing, UK. Picture: Mick Harris.
Little Gull minutus, adult, March 01 2014, Lancing, UK. Picture: Mick Harris. May
 Little Gull minutus, adult, May 15 2014, St Petersburg, Russian Federation. Picture: Ekaterina Papchinskaya.
Little Gull minutus, adult, May 15 2014, St Petersburg, Russian Federation. Picture: Ekaterina Papchinskaya.