 American Herring Gull (smithsonianus)
American Herring Gull (smithsonianus)
(last update: October 30, 2015)
American Herring Gull adult September
IDENTIFYING THE SEX OF MASSACHUSETTS HERRING GULLS BY LINEAR MEASUREMENTS
- DANIEL R. EVANS, PAUL M. CAVANAGH, THOMAS W. FRENCH & BRADFORD G. BLODGET -
IN: J. Field Ornithol., 66(1):128-132 Winter 1995.
Abstract.
Discriminant functions capable of determining sex are lacking for Herring Gull (Larus argentatus) populations in the northeastern United States. A new discriminant function, based on culmen and tarsus lengths, that correctly classified the sex of 94% of gulls collected in Massachusetts was derived. This function is applicable to nesting populations of Massachusetts Herring Gulls, but should be tested against the morphometrics of known-sex gulls before being applied to other populations.

Discriminant functions derived from morphometric data are commonly used to sex a variety of avian species. Within species, geographic variation (e.g., Coulson et al. 1983, Evans et al. 1993, Threlfall and Jewer 1978) and differences between captive and wild populations (e.g., Smith and Wiemeyer 1992) may reduce a function's applicability. For these reasons, functions should be applied only to the populations from which they were derived (e.g., Evans et al. 1993, Smith and Wiemeyer 1992).
Numerous functions have been derived to discriminate between the sexes of Herring Gulls (Larus argentatus). Functions have been derived for populations in Michigan (Shugart 1977), Ontario (Fox et al. 1981), Newfoundland (Coulson et al. 1983), and Great Britain (Coulson et al. 1983, Threlfall and Jewer 1978). As Herring Gulls vary geographically (e.g., Monaghan et al. 1983, Threlfall and Jewer 1978), and may vary clinally between Maine and New York (Threlfall and Jewer 1978), existing discriminant functions may not be applicable to gulls in the northeastern United States.
Roseate Tern (Sterna dougallii) recovery efforts in Massachusetts resuited in the death of a large sample of Herring Gulls from a single breeding colony (Blodget and Henze 1992). This sample provided an opportunity to gather morphological data on the Massachusetts population. Here we present this information and provide a discriminant analysis formula that can accurately sex Massachusetts Herring Gulls.

METHODS
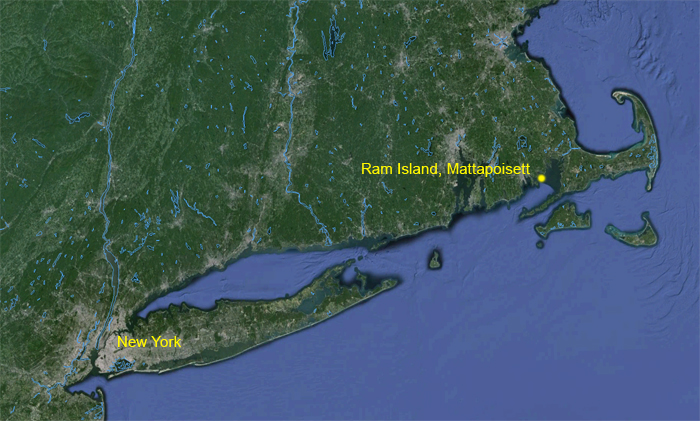
We measured 444 Herring Gulls collected on and around Ram Island, Mattapoisett, Massachusetts (41ø37'N, 70ø48'W) following application of the avicide DRC-1339 on 15 May 1990. All but two gulls are believed to have died as a result of ingesting DRC-1339.
We measured culmen length, tarsus length, wing-chord (flattened) and mass of each gull. All linear measurements were taken on the right side of the body, unless the structure to be measured was absent or damaged. Culmen and tarsus lengths were measured with calipers to the nearest 1 mm, wing-chord was measured with a ruler to the nearest 1 mm, and mass was measured with a 4000-g electronic balance (Metier Instrument Co., Hightstown, New Jersey) to the nearest 0.1 g. Gulls were then dissected and their sexes determined on the basis of internal morphology. Only adult gulls (i.e., those showing full adult plumage) were included in our analysis; this resulted in the exclusion of 22 birds from our data set. We used a two-tailed, two-sample, t-test (SYSTAT, Wilkinson 1991) to test for significant (P < 0.025) intersexual differences in size and mass.
A discriminant function was developed following the guidelines of Frank et al. (1965) as described by Fox et al. (1981) for the V1 validation method. The measured gulls were divided into two samples, the analysis sample and the validation sample. We derived a base discriminant function from all four variables using the MGLH program in SYSTAT (Wilkinson 1991). The combination of measurements that best discriminated between sexes was selected from our base function. We then derived our first discriminant function using the analysisample (n = 316, 75%), and tested the function for accuracy using the validation sample (n = 106, 25%). This process was performed to determine the level of bias in our sample. Once minimal bias was established, a second function was derived by combining analysis and validation samples (n = 422).
| TABLE 1. Measurements of male and female Herring Gulls from Ram Island, Mattapoisett, Massachusetts. | ||||||||||
| . | Males | Females | . | . | ||||||
| Variable | n | Mean ± SD | (range) | CV | n | Mean ± SD | (range) | CV | t | P |
| Culmen (mm) | 169 | 56.0 ± 2.3 | (56-70) | 3.7 | 253 | 57.2 ± 1.9 | (52-62) | 3.4 | -28.5 | 0.001 |
| Tarsus (mm) | 169 | 71.1 ± 2.2 | (65-76) | 3.1 | 253 | 66.0 ± 2.0 | (60-73) | 3.1 | -24.0 | 0.001 |
| Wing chord (mm) | 169 | 429 ± 10.3 | (400-460) | 2.4 | 253 | 408 ± 9.6 | (384-447) | 2.3 | -20.8 | 0.001 |
| Mass (g | 169 | 1147 ± 77.7 | (973-1143) | 6.8 | 246 | 1023 ± 106 | (718-1385) | 10.4 | -13.7 | 0.001 |
RESULTS
Massachusetts Herring Gulls exhibited significant inter- and little intrasexual morphological variation. Males were significantly larger than females for all measurements (P = 0.001, Table 1). Culmen, tarsus and wing-chord lengths varied little within sexes (CV = 2.3-3.7%), whereas mass was more variable (CV -- 6.8-10.4%, Table 1).
Herring Gulls could be reliably sexed using only two measurements. From our base discriminant function, the combined measurements of culmen (CL) and tarsus (TL) lengths were the most discriminating factors in classifying the sex of Herring Gulls (Wilk's lambda = 0.261, X 2 = 418.836, df = 2, P < 0.001). The classification function:
(CL X 0.853) + (TL X 0.736) = 101.843
correctly classified 95% of the Herring Gulls in the analysis sample. If the function value was greater than 101.843, the gull was classified as a male; gulls with values less than or equal to the function value were classified as females. This function correctly classified 187 of 192 females (97%) and 113 of 124 males (91%). Using the validation sample, the function correctly classified 56 of 61 females (92%), and 42 of 45 males (93%), providing an overall accuracy of 92%. The similarity of our classification accuracy derived from the analysis sample (n = 316, 95%) to that derived from the validation sample (n = 106, 92%) suggests minimal sampling bias. Our second classification function, derived from combined analysis and validation samples, also accurately discriminated between the sexes (Wilk's lambda = 0.273, X 2 = 542.634, df = 2, P < 0.001). The classification function:
(CL X 0.882) + (TL X 0.730) = 103.178
correctly classified 94% of the Herring Gulls. This function should be used when sexing Herring Gulls from Massachusetts because of its high accuracy and minimal bias, due to its larger sample size.

DISCUSSION
Body-mass values for Massachusetts gulls were influenced by the use of avicide. Decino et al. (1966) reported that European Starlings (Sturnus vulgaris) poisoned with DRC-1339 doubled their water consumption 4-8 h after ingestion and experienced hemorrhaging and congestion in their kidneys that reduced their ability to excrete toxic compounds. Additionally, they reported fat-like accumulations of uric acid in the body cavity.
We observed similar accumulations, as well as accumulations of fluids, while dissecting Massachusetts gulls and believe these materials affected our body-mass values. Therefore, masses reported in this study are not directly comparable to values reported in previous studies (i.e., those involving non-poisoned gulls), nor were they considered for incorporation into our discriminant function.
The accuracy of our discriminant function was similar to those reported for other populations of Herring Gulls. Shugart (1977) reported that a discriminant function based on combined head and bill length, and wingchord, correctly sexed 98.15% of the Michigan Herring Gulls tested. A function based on seven variables (wing length, mass, tarsus length, culmen depth, culmen length, tail length, and the product of culmen depth by length) correctly identified the sex of 91.3% of British and 97.7% of Newfoundland gulls tested (Threlfall and Jewer 1978). Fox et al. (1981) derived two functions to determine the sex of Herring Gulls in Ontario. The first function, based on head and foot lengths, correctly identified the sex of 94.9% of gulls tested; the second function, based on head length and bill depth, correctly identified the sex of 96.5% of gulls tested. Coulson et al. (1983) reported accuracies of 95.8-96.7%, depending on whether one (combined head and bill length) or five (combined head and bill length, body-mass, bill depth, bill length, and wing length) variables were used to derive the function. Our function's accuracy falls within the range of values reported previously. Although based on different variables, the high levels of accuracy reported in these studies suggest identifiable sexual differences in morphology exist throughout the Herring Gull's range. Determination of which function is appropriate depends on the population under investigation.
Our discriminant function was developed from, and intended to be applied to, nesting populations of Massachusetts Herring Gulls. We recommend that those considering applying this function to other populations first test the morphometrics of known sex gulls against our equation (e.g., Evans et al. 1993) to determine if it can correctly classify birds from other populations.
ACKNOWLEDGMENTS
Roseate Tern restoration activities on Ram Island were funded by the Massachusetts Division of Fisheries and Wildlife (MDFW) with partial support from the U.S. Fish and Wildlife Service under Section 6 of the Endangered Species Act. We thank J. E. Cardoza, W. J. Davis and S. Langlois of the MDFW for assistance in processing gulls. W. Threlfall, K. Yasukawa and an anonymous reviewer provided helpful comments on this manuscript.
LITERATURE CITED
See PDF.
Great Lakes birds
 American Herring Gull (smithsonianus) 0846-10957 30th cycle (30CY), September 20 2015, New Buffalo, Berrien County, Michigan. Picture: Amar Ayyash.
American Herring Gull (smithsonianus) 0846-10957 30th cycle (30CY), September 20 2015, New Buffalo, Berrien County, Michigan. Picture: Amar Ayyash. American Herring Gull (smithsonianus) 0966-44575 18th cycle (18CY), September 15 2016, New Buffalo, MI. Picture: Amar Ayyash.
American Herring Gull (smithsonianus) 0966-44575 18th cycle (18CY), September 15 2016, New Buffalo, MI. Picture: Amar Ayyash. American Herring Gull (smithsonianus) 1106-03794 6CY, September 07 2015, New Buffalo, Berrien County, Michigan. Picture: Amar Ayyash.
American Herring Gull (smithsonianus) 1106-03794 6CY, September 07 2015, New Buffalo, Berrien County, Michigan. Picture: Amar Ayyash. American Herring Gull (smithsonianus) 1106-08886 9th cycle (9CY), September 15 2016, New Buffalo, MI. Picture: Amar Ayyash.
American Herring Gull (smithsonianus) 1106-08886 9th cycle (9CY), September 15 2016, New Buffalo, MI. Picture: Amar Ayyash. American Herring Gull (smithsonianus) 1106-13326 7th cycle (7CY), September 20 2015, New Buffalo, Berrien County, Michigan. Picture: Amar Ayyash.
American Herring Gull (smithsonianus) 1106-13326 7th cycle (7CY), September 20 2015, New Buffalo, Berrien County, Michigan. Picture: Amar Ayyash. American Herring Gull (smithsonianus) 1106-13485 7th cycle (7CY), September 20 2015, New Buffalo, Berrien County, Michigan. Picture: Amar Ayyash.
American Herring Gull (smithsonianus) 1106-13485 7th cycle (7CY), September 20 2015, New Buffalo, Berrien County, Michigan. Picture: Amar Ayyash. American Herring Gull (smithsonianus) 1116-38317 9CY, September 07 2015, New Buffalo, Berrien County, Michigan. Picture: Amar Ayyash.
American Herring Gull (smithsonianus) 1116-38317 9CY, September 07 2015, New Buffalo, Berrien County, Michigan. Picture: Amar Ayyash.East Coast birds
 American Herring Gull (smithsonianus) adult AAR September 09 2011, New London, Prince Edward Island.
American Herring Gull (smithsonianus) adult AAR September 09 2011, New London, Prince Edward Island. American Herring Gull (smithsonianus) adult F01 May 2009 & September 2010, New Hampshire. Picture: Kathy Abbott & Bill Clark.
American Herring Gull (smithsonianus) adult F01 May 2009 & September 2010, New Hampshire. Picture: Kathy Abbott & Bill Clark. American Herring Gull (smithsonianus) K30 2CY & 6CY, August 2011 & July-September 2015, Hampton Beach, NH. Picture: Gustine Silva-Burke, Robbie & Colleen Prieto.
American Herring Gull (smithsonianus) K30 2CY & 6CY, August 2011 & July-September 2015, Hampton Beach, NH. Picture: Gustine Silva-Burke, Robbie & Colleen Prieto. American Herring Gull (smithsonianus) adult K68 May 2011, February & March 2012 & September 2013, Hampton Beach, NH.
American Herring Gull (smithsonianus) adult K68 May 2011, February & March 2012 & September 2013, Hampton Beach, NH. American Herring Gull (smithsonianus) adult M37 September 28 2013, Hampton Beach, NH. Picture: Jon Worthen.
American Herring Gull (smithsonianus) adult M37 September 28 2013, Hampton Beach, NH. Picture: Jon Worthen. American Herring Gull (smithsonianus)7cy-8cy M54 (formerly C45) 2011, 2012 & 2013, Hampton Beach, NH. Picture: Keith Mueller & Jon Worthen.American Herring Gull (smithsonianus) T47 adult, April 2012, September 2013 & September 2015, Odiorne Point, New Hampshire Coast.
American Herring Gull (smithsonianus)7cy-8cy M54 (formerly C45) 2011, 2012 & 2013, Hampton Beach, NH. Picture: Keith Mueller & Jon Worthen.American Herring Gull (smithsonianus) T47 adult, April 2012, September 2013 & September 2015, Odiorne Point, New Hampshire Coast. American Herring Gull (smithsonianus) 4J adult, September 2016 & 2018, November 2017, Quidi Vidi Lake - St. John's, Newfoundland, Canada. Picture: Lancy Cheng.
American Herring Gull (smithsonianus) 4J adult, September 2016 & 2018, November 2017, Quidi Vidi Lake - St. John's, Newfoundland, Canada. Picture: Lancy Cheng. American Herring Gull (smithsonianus) XS adult, September 25 2018, Quidi Vidi Lake - St. John's, Newfoundland, Canada. Picture: Lancy Cheng.
American Herring Gull (smithsonianus) XS adult, September 25 2018, Quidi Vidi Lake - St. John's, Newfoundland, Canada. Picture: Lancy Cheng. American Herring Gull (smithsonianus) Q Q Q Fed band Q Q adult, September 05 2009, Presqu'ile, Brighton, ON. Picture: Newfoundlander61.
American Herring Gull (smithsonianus) Q Q Q Fed band Q Q adult, September 05 2009, Presqu'ile, Brighton, ON. Picture: Newfoundlander61. American Herring Gull (smithsonianus) adult, September 22 2012, Plum Island, Massachusetts, USA. Picture: Suzanne Sullivan.
American Herring Gull (smithsonianus) adult, September 22 2012, Plum Island, Massachusetts, USA. Picture: Suzanne Sullivan. American Herring Gull (smithsonianus) adult, September 13 2011, Acadia National Park Maine, USA. Picture:Thomas Gass.
American Herring Gull (smithsonianus) adult, September 13 2011, Acadia National Park Maine, USA. Picture:Thomas Gass.