 Larus cachinnans
Larus cachinnans
(last update:
Greg Neubauer
Marcin Przymencki
Albert de Jong
Mars Muusse
 sub-adult cachinnans: March
sub-adult cachinnans: March
In 2011, Chris Gibbins, Grzegorz Neubauer and Brian Small published two extensive papers in Britsih Birds, dealing with Caspian Gull. Below, you will find the content of the second paper "phenotypic variability and the field characteristics of hybrids".
The full title reads: From the Rarities Committee's files - Identification of Caspian Gull. Part 2: phenotypic variability and the field characteristics of hybrids, by Chris Gibbins, Grzegorz Neubauer and Brian Small, IN: BB 104/2011. ORDER PAPER COPY!
"we" in the text below refers to the original authors. If any errors occur in this text, please let me know and mail to marsmuusseatgmaildotcom.
PART 1: ABSTRACT & INTRODUCTION
PART 2: APPROACH
BELOW: PART 3 OF THE PAPER
Identification of Caspian Gull. Part 2: phenotypic variability and the field characteristics of hybrids
Adult birds
 Larus cachinnans 10CY female 17P1 April 30 2009, Włocławek Reservoir, central Poland. Picture: Magdalena Zagalska-Neubauer. Ringed as chick in 2000 within the core range in southern Poland. Note near thayeri-pattern on P10. |
 Larus cachinnans 10CY male 802P April 29 2006, Włocławek Reservoir, central Poland. Picture: Magdalena Zagalska-Neubauer. Ringed as chick in 1997. |
Adults were scored using measurements, and the appearance of features observed in photographs of the head and wing-tip of birds captured in Ukrainian and Polish colonies during the breeding season (Caspian n=100, Herring n=132, known hybrids n=12). All the adult Herring Gulls sampled were from Poland, and so were of the nominate race argentatus. Birds of this race can be extremely similar to Caspian Gull in plumage (e.g. primary pattern) and bare-part colours, and thus are a potential pitfall. It was not felt necessary to include L. a. argenteus in the sample because this race differs considerably from Caspian in adult plumage and so should not present any identification problems.
 Eye pigmentation and orbital-ring colour were assessed in the field, with each assigned a score as per table 1. Wings were held open, photographed and a number of aspects of the primary pattern were assessed from these images. Advances in digital photography mean that it is possible to capture high-quality images of birds in the field, either in flight or while wing-stretching, and these can be used by birders to assess details of the wing-tip pattern. Thus, while the scoring system is relatively detailed, it should be possible to capture all the necessary features in photographs of birds in the field and hence score an adult gull’s wing-tip pattern in the same way as we have done using photographs of birds in the hand.
Eye pigmentation and orbital-ring colour were assessed in the field, with each assigned a score as per table 1. Wings were held open, photographed and a number of aspects of the primary pattern were assessed from these images. Advances in digital photography mean that it is possible to capture high-quality images of birds in the field, either in flight or while wing-stretching, and these can be used by birders to assess details of the wing-tip pattern. Thus, while the scoring system is relatively detailed, it should be possible to capture all the necessary features in photographs of birds in the field and hence score an adult gull’s wing-tip pattern in the same way as we have done using photographs of birds in the hand.
Bill shape was expressed as the ratio of bill length (measured in the standard way, along the culmen) to depth at the gonys, with the ratio being scored as per table 1. Leg length was measured in millimetres and then converted to a trait score, to reflect how long-legged each bird might appear in the field. The trait score for each bird was based on its length relative to the mean length of all individuals of that sex in the sample; since we were interested in the relative leg length of an individual irrespective of species, the means were calculated from data pooled across both species (but separately for each sex). Thus, each bird was scored as having relatively short, relatively long or average-length legs, given its sex. We did not include leg colour in the adult criteria as this is well known to overlap completely between Herring and Caspian Gull - both species can have either pink or bright yellow legs and so this feature is of little value in identification (details in part 1).
First-winter birds
For 1W birds, traits included aspects of structure, plumage and moult (table 2). Scoring of sample birds (Herring n=85, Caspian n=63, hybrids n=12) was undertaken using digital photographs taken in the field between October and March. The 1W Herring Gulls were all from Scotland, while the Caspian Gulls were from the Ukraine, Romania, Poland and Latvia. Only perfectly side-on images were used, because of the need to measure relative lengths and depths (and calculate ratios) for bill and wing-tip features.

Larus cachinnans 2cy, 14 March 2012, Texel, the Netherlands. Picture: Herman Bouman. Picture, with arrows to explain moult to 2nd gen feathers ( click image for details). Scorings as per table 2.
The moult traits represented an assessment of the extent of post-juvenile moult. This moult takes place soon after first-calendar-year (1CY) individuals gain independence and continues through the autumn, before being suspended for the winter. To score birds it was first necessary to set a cut-off point that represents the end of the moult. We did this based on moult scores for a sample of Caspian Gulls (fig. 2). The maximum scores were reached by October, with no statistically significant increase beyond this point. This indicates that most birds complete the post-juvenile moult by October, with no (or only exceptional) further replacement of feathers beyond this point. Thus, to assess the final extent of the post-juvenile moult, sample birds had to be from October or later. Moult commences again in the spring, with feathers being dropped and replaced from April onwards. We therefore set the window within which to sample 1W birds as being from October to March inclusive.
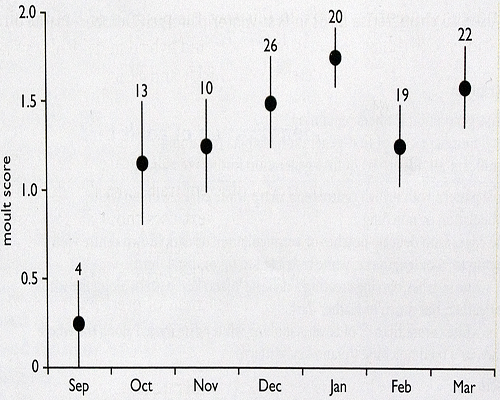
Fig. 2. Moult scores (mean and SE) from a sample of first-winter Caspian Gulls Larus cachinnans in southern Poland (n = 114).The score for each bird is the sum of values for moult in its median and greater coverts and tertials.
For coverts, scores are categorical:
0 = no moult,
1 = 1-3 new feathers,
2 = more than 3 feathers but less than a third of total,
3 = between a third and a half of all feathers, and
4 = more than half of all feathers in the group.
Tertial scores are simply the number of new feathers present. Values shown in the plot are the number of individuals scored in each month.
Primary projection (fig. 3) and the presence of a ventral bulge are potentially useful in identification, and so were assessed for 1W birds. However, differences in posture mean that it is not possible to measure and assess these traits in the hand in a way that is useful for field identification. Thus, primary projection and presence of a ventral bulge were not assessed for adult birds, and we assume that results for 1W birds are valid for adults.
 |
 |
| Fig. 3. Measurements used to derive bill-length to depth and primary-projection ratios for the sample of first-winter birds. Bill ratio is calculated as L/D, primary projection as I/L (projection beyond tail/length beyond tertials). Both images show same juvenile Caspian Gull Larus cachinnans, August 23 2008, IJmuiden, the Netherlands. Picture: Mars Muusse. | |
The full length of the culmen is not visible from the side because of feathering. Thus, for 1W birds we had to use a different bill-length dimension from that for adults. We assessed bill length in 1W birds from the distal point of the feathering to the bill tip and used this to calculate length/depth ratios and scores (see fig. 3). We do not use these data to generate overall bill length/depth frequency statistics for Caspian versus Herring Gull (we use the adult data for this), but instead use them to calculate overall trait scores for individual 1W birds (see “Data analysis”). The different measuring techniques result in different ratio values (greater for in-hand measurements), but the relative differences between species remain constant (see Neubauer et al. 2006 for discussion). Appendix I shows the raw ratios calculated for all 1W birds from photographs; these should be useful for birders assessing ratios in the field.
Relative leg length for 1W birds was assessed from photographs, with each bird scored according to whether its legs looked short, moderately long or long. As with the bill-length dimensions, these data were used only to calculate sum trait scores for each bird; assessment of the frequency of different leg lengths in the two species is based only on data for adults.
In very few cases were high-quality photographs available both in flight and on the ground for the same individual gull. While traits visible only in flight are important for field identification of 1W birds (specifically the underwing pattern), the lack of matching image sets meant that it was not possible to incorporate in-flight criteria into the scoring system for this age group. We therefore treat variability in the underwing pattern separately.
END OF PART 3
CONTINUE PART 4: DATA ANALYSIS & IDENTIFICATION OF ADULT BIRDS
 Larus cachinnans 1CY & 4CY UKK L-006940 September & October 2010, Denmark & March 2013, the Netherlands.
Larus cachinnans 1CY & 4CY UKK L-006940 September & October 2010, Denmark & March 2013, the Netherlands. Larus cachinnans sub-adult UKK xxx March 06 2011, Wörth, Austria. Picture: Wolfgang Schweighofer.
Larus cachinnans sub-adult UKK xxx March 06 2011, Wörth, Austria. Picture: Wolfgang Schweighofer.  Larus cachinnans 2CY-4CY PACX September 2010 - March 2012, Deponie Pohlsche Heide - Minden, Germany (52°23'05N, 08°46'45E).
Picture: Armin Deutsch.
Larus cachinnans 2CY-4CY PACX September 2010 - March 2012, Deponie Pohlsche Heide - Minden, Germany (52°23'05N, 08°46'45E).
Picture: Armin Deutsch. Larus cachinnans 4CY PDNP January & March 2013, Switzerland. Picture: Julien Torre & Laurent Valloton.
Larus cachinnans 4CY PDNP January & March 2013, Switzerland. Picture: Julien Torre & Laurent Valloton. Larus cachinnans PHLD 3CY-4CY, March - July 2013 & March 2014, CET Champteussé-sur-Baconne, France. Picture: Alain Fossé & Olivier Laluque.
Larus cachinnans PHLD 3CY-4CY, March - July 2013 & March 2014, CET Champteussé-sur-Baconne, France. Picture: Alain Fossé & Olivier Laluque. Larus cachinnans 4CY 7P02 May 17-23 2007, Łubna landfill, Poland. Picture: Ruud Altenburg.
Larus cachinnans 4CY 7P02 May 17-23 2007, Łubna landfill, Poland. Picture: Ruud Altenburg.  Larus cachinnans 3CY-5CY PLG DN-03919 January 2006 - November 2008, Deponie Pohlsche Heide - Minden, Germany (52°23'05N, 08°46'45E).
Larus cachinnans 3CY-5CY PLG DN-03919 January 2006 - November 2008, Deponie Pohlsche Heide - Minden, Germany (52°23'05N, 08°46'45E). Picture: Armin Deutsch.
 Larus cachinnans A+ 2CY & 5CY, February 13 2013 & March 02 2016, Amsterdam & Barneveld waste dump, the Netherlands.
Larus cachinnans A+ 2CY & 5CY, February 13 2013 & March 02 2016, Amsterdam & Barneveld waste dump, the Netherlands.
Larus cachinnans 4CY, March 28 2013, Roermond, the Netherlands. Picture: Theo Muusse.

Larus cachinnans 4CY, March 28 2013, Roermond, the Netherlands. Picture: Theo Muusse.